Памяти Нины Николаевны Протасовой
18 июня 2005года не стало Нины Николаевны Протасовой известного физиолога растений, одного из ведущих специалистов в области светокультуры растений.
Нина Николаевна родилась 30 ноября 1922 года в семье служащих, после школы она поступила в Тимирязевскую сельскохозяйственную академию, которую успешно окончила в 1946 году. Затем она работала в системе Министерства сельского хозяйства и одновременно занималась научной работой. В 1949 году она поступила в очную аспирантуру Института физиологии растений им. К.А.Тимирязева АН СССР, с которой затем уже не расставалась никогда.
Для своей научной работы Нина Николаевна избрала новое перспективное направление - светокультуру растений, которому она оставалась верной всю свою жизнь. В своих исследованиях она придавала большое значение разработке приёмов выращивания растений, имеющих большое хозяйственное значение. Нина
Николаевна успешно защитила диссертацию на соискание кандидата биологических наук по теме: "Применение искусственного досвечивания при выращивании рассады огурцов и томатов".
Нина Николаевна внесла важный вклад в изучение физиологических особенностей выращивания растений в условиях защищенного грунта с использованием искусственного света как в условиях досвечивания, так и при полностью искусственном свете. Эта работа имела большое значение для продвижения промышленной
светокультуры в северные районы страны. Ценными были и выводы о физиологической оценке эффективности излучения ламп. Много сил и времени потратила она при выработке требований к техническим характеристикам новых источников света, их биологическим испытаниям и последующему внедрению в практику промышленной светокультуры. С её именем связано внедрение в светокультуру трубчатых ксеноновых ламп в 60-70 годы, трёхфазных металлогалогенных ламп в 70-80 годы. Она опубликовала более 60 научных работ, получила несколько авторских свидетельств, её трижды награждали медалью на ВДНХ СССР за внедрение научных разработок в тепличное растениеводство. Много раз
Н. Н. Протасова возглавляла межведомственные и государственные аттестационные комиссии по приемке новых ламп для растениеводства, участвовала в работе научных конференций, её приглашали для чтения лекций на методических сборах для работников сельского хозяйства.
Для всех, кто её знал или работал с нею, Н. Н. Протасова навсегда осталась очень доброжелательным, оптимистичным, неравнодушным и веселым человеком, знающим и отзывчивым, готовым поделиться своими знаниями и опытом специалистом. Она по праву стоит в одном ряду с такими пионерами светокультуры в нашей стране, как академик Н. А. Максимов, член кор. А. А. Ничипорович, чл. кор. Б. А. Мошков, профессор А.Ф. Клешнин, профессор В.М. Леман, доктор биологических наук Н. П. Воскресенская с которыми ей посчастливилось не только быть знакомой , но и работать.
Выпускница Тимирязевской академии, заслуженный светофизиолог
Нина Николаевна Протасова
(1922 – 2005)
«Мир Теплиц» №6/05
30 ноября 2002 г. отмечалось 80-летие Нины Николаевны Протасовой - ведущего специалиста в
области светофизиологии и светокультуры растений, старшего научного сотрудника Института физиологии растений РАН.
Очень скоро после окончания Тимирязевской сельскохозяйственной академии (ТСХА) в 1946г, приоритетной для молодого
специалиста стала научная работа и уже в 1949г, К.Н.Протасова поступила в очкую аспирантуру при ИФР АН СССР. С тех
пор ее жизнь неразрывно связана с этим институтом.
Она успешно защитила диссертацию по теме "Применение искусственного досвечивания при выращивании рассады огурцов
и томатов - в те годы это была новаторская работа, не утратившая своей актуальности и в наши дни.
Работая бок о бок с такими выдающимися отечественными светофизиологами растений, как А.А.Ничипорович, А.Ф.Клешнин,
Н.П.Воскресенская, Нина Николаевна внесла большой вклад не только в разработку физиологических основ интенсивной
светокультуры но и в формирование современных представлений о характеристиках эффективности оптического излучения
для выращивания высших растений в защищенном грунте.
Творческий диапазон Н.Н.Протасовой как исследователя был необычайно широк; об этом свидетельствуют выполненные
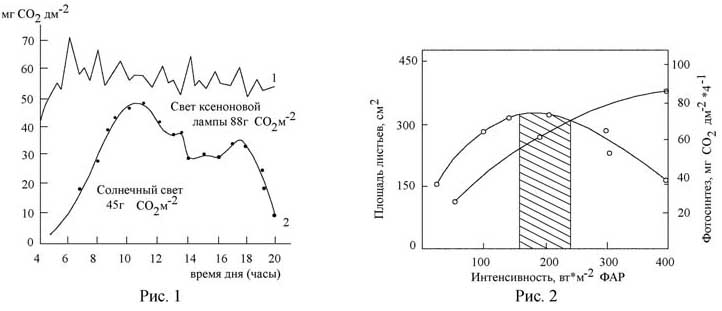
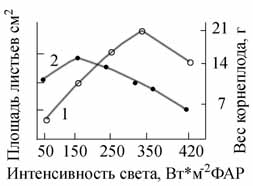
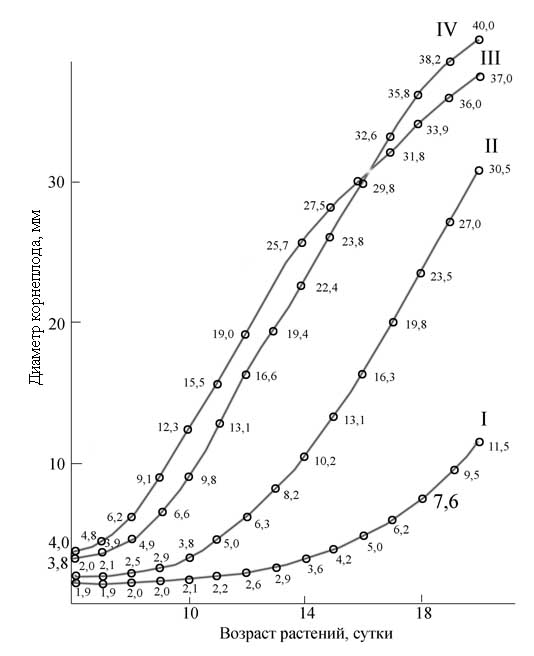
ею в разные годы значительные работы по изучению влияния спектрального состава оптического излучения на фотосинтез
растений с использованием, в т.ч., метода "световых кривых", и эксперименты, установившие решающее значение фактора,
интенсивности света при выращивании сельскохозяйственный растений в искусственных условиях, а также исследования
фитофизиологической значимости различных участков ФАР, УФ области спектра и др.
Особо следует отметить постоянный интерес Н.Н.Протасовой к проблеме обоснования и внедрения
в практическое растениеводство новых эффективных газоразрядных источников излучения. После длительных и детальных
биологических испытаний в 60-е годы получили "путевку в жизнь" трубчатые ксеноновые лампы, и в 70-80-е годы - трехфазные МГЛ.
Неоднократно К.Н.Протасова возглавляла межведомственные и государственные аттестационные комиссии по новым
растениеводческим лампам.
Нина Николаевна - автор более 60 научных работ, а также нескольких авторских свидетельств.
За внедрение достижений биологической науки в тепличное растениеводство она трижды удостаивалась медалей ВДНХ СССР,
Мы помним ее как истинно интеллигентного высококультурного, доброжелательного человека знающего и отзывчивого специалиста,
готового поделиться своим опытом и знаниями. Такой ее знают все, кому пришлось с ней работать.
Copyright © ООО "Воля"